Most models of behavior change assume that people continue habits despite the harm they cause because they cannot visualize the consequences, and if one were simply to understand the risks clearly enough, behavior would change. In practice, this explanation fails to capture the key paradox of addiction and behavioral health: people often continue to harm themselves even with reasoning perfectly intact.
Consider two groups that at first glance appear quite different. On one end, there are high-functioning substance users, often maintaining careers, relationships, and perfectly normal daily routines. The substance use is sprinkled in subtly enough that their lives remain stable, and the negative feedback normally associated with addiction never fully materializes. Income, structure, and external success buffer them from the consequences that might otherwise trigger behavioral change (Kelly et al., 2017; Witkiewitz et al., 2019).
Situated on the other end of the spectrum are individuals in the pre-contemplative stage of change, who may experience more instability, but regardless, do not yet believe their behavior to be a problem. The habit thus persists because it does not seem harmful enough to demand the user’s attention. Their internal signals that normally indicate misalignment, stress, and discomfort are faint and unclear.
These two populations appear very different to an outsider. One maintains an image of control, the other comes across as disengaged or unaware. Yet, in both cases, the underlying mechanism is the same: the brain never receives a signal strong enough to reevaluate behavior.
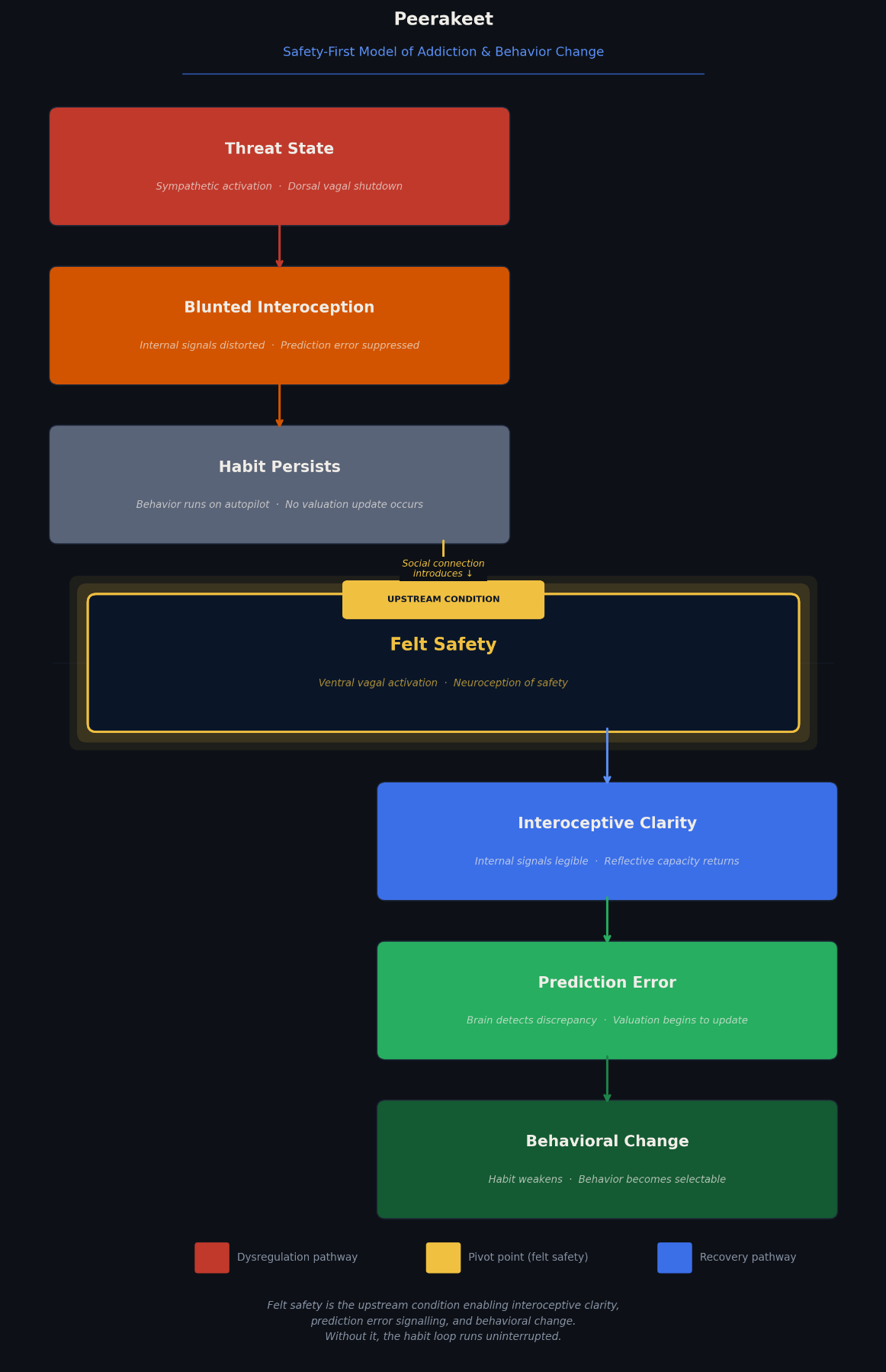
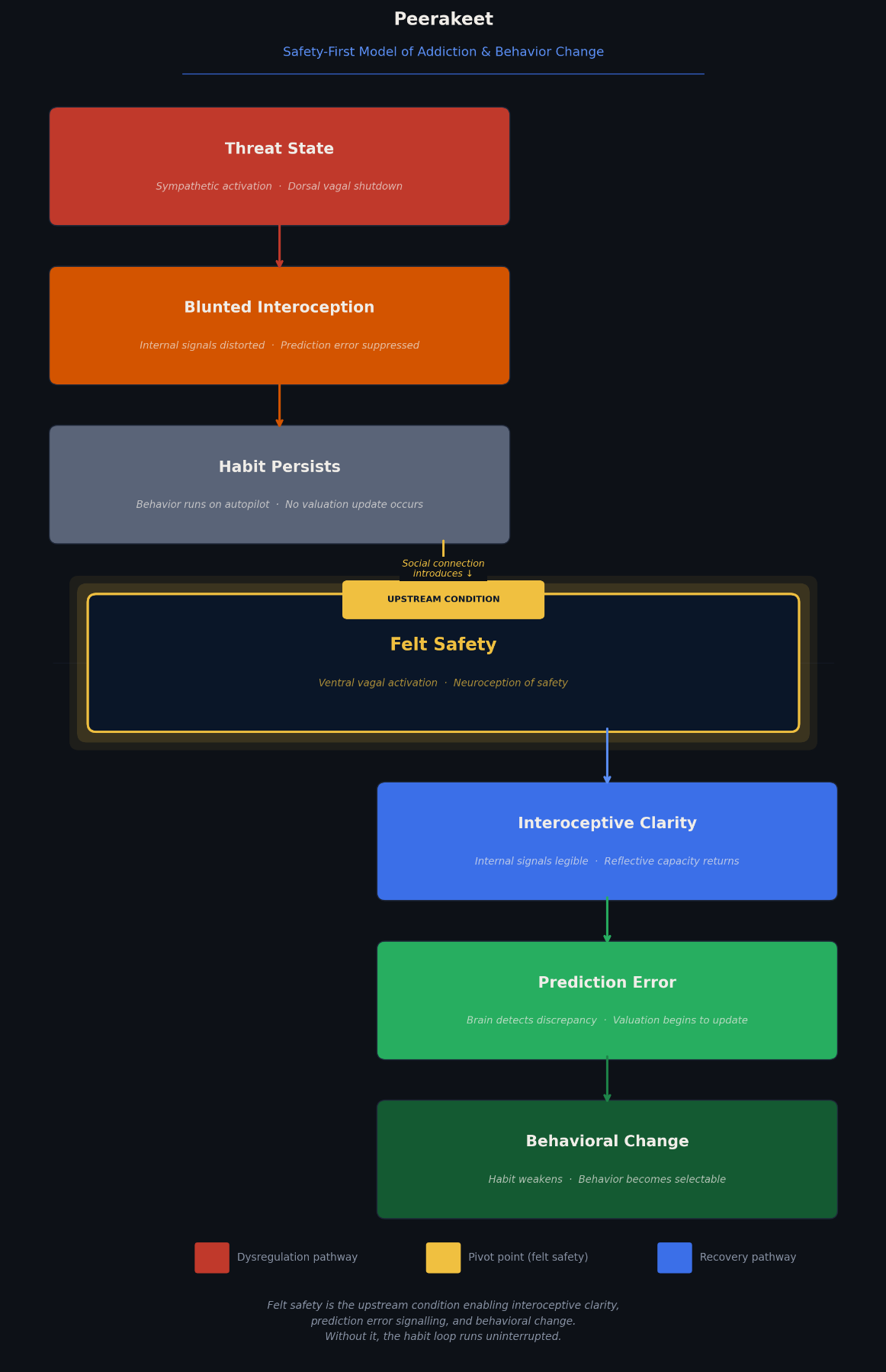
Without this signal, habits run on autopilot.
The Habit Loop and Why it Persists
Habits form through a neurological cycle: environmental cues trigger a behavior, the behavior produces a reward, and the brain updates the expected value of the cue (Everitt & Robbins, 2016). Over time, dopamine activity shifts away from the reward itself and towards the cue that predicts it (Schultz, 1997). Once this shift occurs, behavior becomes automatic.
It is at this stage that the habit no longer depends on how pleasurable the reward actually feels. The brain runs the behavior based on prediction rather than experience. This is how habits often persist even in the face of individuals who recognize their downsides.
To interrupt this cycle, the brain must detect a discrepancy between the expected reward and the actual outcome, a signal known as a prediction error. When the outcome is worse than expected, the brain gradually updates its valuation of the cue and the habit weakens (Schultz, 2016).
But prediction errors only occur when the brain clearly feels the outcome, and that depends on something else entirely.
Under conditions of stress or threat, it has been shown that internal signals become distorted, blunted by the emotional noise (Paulus & Stein, 2010; Schulz & Vögele, 2015). When facing this moment, individuals may understand a behavior intellectually while remaining unable to feel its consequences.
If the sensory system cannot even flag the discrepancy, then the valuation system of the brain will have nothing to update. Before prediction error, before insight, before motivation to enact change, there must be a shift in bodily state toward what is described as felt safety.
Felt safety is the upstream condition that allows internal signals to become clear, reflective systems to reengage, and valuation to update. Once safety is established, interoceptive precision improves, reflective capacity returns, cues lose strength, and behavior becomes selectable (Paulus & Stein, 2010; Schulz & Vögele, 2015). This concept draws heavily from the work of Stephen Porges and polyvagal theory, which proposes that the nervous system constantly evaluates environmental cues for safety or threat (Porges, 2011). When safety is detected, the “social engagement system” becomes available, enabling reflection, connection, and flexible behavior (Porges, 2011). While polyvagal theory has been clinically influential in understanding how felt safety regulates the nervous system, some scholars have noted that its mechanisms are more complex and context-dependent than originally proposed, highlighting the need to treat it as an organizing model rather than a concrete anatomical account (Grossman & Taylor, 2007).
This framework explains why two seemingly opposite populations, high-functioning users and pre-contemplators, respond to connection more than confrontation. It suggests that habit change is not prompted by information or punishment, but by a state of safety that allows the brain to feel what it has been ignoring.
The question then becomes: what actually produces that state? The answer, consistently across the literature, is other people.
Creating Conditions of Safety Through Social Connection
Social connection has been demonstrated to regulate the nervous system. The presence of a trusted person reduces activation of the body’s mounted stress response, lowers cortisol levels, and dampens threat signaling (Hostinar et al., 2014). This shift decreases the metabolic burden that arises from a state of constant vigilance, allowing the brain to operate at a calmer baseline (Coan & Sbarra, 2015).
Connection also activates endogenous opioid pathways associated with bonding and reward, reinforcing feelings of safety and belonging in the ventral striatum, septal area, and amygdala (Inagaki, 2018). Over time, individuals embedded in supportive social networks demonstrate better mental health outcomes, increased resilience, and improved physical health (Holt-Lunstad, 2021).
However, social connection does not universally produce safety. In some contexts, interpersonal connections can activate threat responses, shame, or hypervigilance, particularly in populations where relationships themselves have historically been sources of harm (Hostinar et al., 2014; Holt-Lunstad, 2021). This introduces an important distinction between social presence and relational safety: having people around is not the same as having connections that regulate the nervous system, and social proximity alone does not guarantee physiological regulation. It is the quality and perceived safety of the relationship that determines whether the nervous system downregulates threat signaling (Coan & Sbarra, 2015; Hostinar et al., 2014).
A person surrounded by co-users in a high-stress environment has social contact without felt safety. Research on social contagion and behavioral networks shows that substance use patterns can spread through social ties, particularly in environments where substance use becomes normative (Christakis & Fowler, 2008; Kelly et al., 2017). In these environments, nervous system dysregulation is common: high stress, low trust, chronic threat. The peer influence effect is therefore not solely about behavioral modeling, but also about the physiological state the environment maintains. Without neuroception of safety, the brain cannot fully access the social engagement system (Porges, 2011). In that state, interoceptive clarity is reduced, reflection becomes harder, and habits continue operating automatically.
Research on loneliness offers a mirror image of the argument. Chronic isolation elevates threat vigilance, increases cortisol baseline, and reinforces stress physiology over time, rendering behavioral reflection difficult (Holt-Lunstad, 2021; Cacioppo & Cacioppo, 2018). The absence of connection is therefore not neutral. It can actively dysregulate the nervous system, meaning individuals without supportive networks begin from a worse neurological baseline for change.
In simpler words, social connection fosters safety. It is this safety that restores the mind’s ability to interpret what the body is communicating. Clear internal signals make reflection possible, allowing the brain to feel costs, evaluate urges, and assign value accurately. Without interoceptive precision, awareness remains intellectual rather than embodied. Insight may exist, but it cannot influence behavior.
This matters for both of our characters, the high-functioning and the pre-contemplative. At both ends of the spectrum, we see subtle costs finally become perceptible. High-functioning individuals may begin to feel fatigue, tension, craving rebound, or stress that had been blurred by buffering. Pre-contemplators may notice discomforts they never registered. Internal signals are now tolerable rather than overwhelming. The reward of the habit starts to feel less comforting and less rewarding. Awareness gains texture.
Why This Matters
This framework suggests that behavior change may begin earlier in the process than many traditional interventions assume. Rather than starting with information or motivation, effective change may start with establishing the correct physiological state. Safety must come first.
It is important to emphasize that the existing evidence largely shows social connection, physiological safety, and improved health outcomes rather than a definitive causal link (Holt-Lunstad, 2021; Hostinar et al., 2014). The argument at stake is not that safety has been definitively proven as the sole prerequisite for change, but that it may function as an upstream condition that enables insight, reflection, and valuation updates to occur.
At Peerakeet, we believe in the power of peer support to create conditions of safety. Without the right state of mind, shared experience and relational trust will have no impact on awareness-based strategies such as reflection, motivational interviewing, or cognitive reframing. If the brain cannot process internal signals clearly, insight will not translate into meaningful change.
Rather than simply motivating individuals, peer relationships may just activate the biological processes required for awareness, reflection, and value updates. This is why the role of peer support is more to us than just an auxiliary feature of behavioral health systems. It is instead the foundational condition through which the neurological prerequisites for habit change become accessible.
The Peerakeet Safety-First Model of Addiction and Behavior Change
[
{kind=link}
References
Cacioppo, J. T., & Cacioppo, S. (2018). Loneliness in the modern age: An evolutionary theory of loneliness. Advances in Experimental Social Psychology, 58, 127–197.
Christakis, N. A., & Fowler, J. H. (2008). The collective dynamics of smoking in a large social network. New England Journal of Medicine, 358(21), 2249–2258.
Coan, J. A., & Sbarra, D. A. (2015). Social baseline theory: The social regulation of risk and effort. Current Opinion in Psychology, 1, 87–91.
Everitt, B. J., & Robbins, T. W. (2016). Drug addiction: Updating actions to habits to compulsions ten years on. Annual Review of Psychology, 67, 23–50.
Grossman, P., & Taylor, E. W. (2007). Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone and biobehavioral functions. Biological Psychology, 74(2), 263–285.
Holt-Lunstad, J. (2021). The Major Health Implications of Social Connection. Current Directions in Psychological Science, 30(3), 251-259.
Hostinar, C. E., Sullivan, R. M., & Gunnar, M. R. (2014). Psychobiological mechanisms underlying the social buffering of the hypothalamic-pituitary-adrenocortical axis: A review of animal models and human studies across development. Psychological Bulletin, 140(1), 256–282.
Inagaki, T. K. (2018). Neural mechanisms of the link between giving social support and health. Annals of the New York Academy of Sciences, 1428(1), 33–50.
Kelly, J. F., Bergman, B. G., Hoeppner, B. B., Vilsaint, C. L., & White, W. L. (2017). Prevalence and pathways of recovery from drug and alcohol problems in the United States population. Drug and Alcohol Dependence, 181, 162–169.
Paulus, M. P., & Stein, M. B. (2010). Interoception in anxiety and depression. Brain Structure and Function, 214(5–6), 451–463.
Porges, S. W. (2011). The polyvagal theory: Neurophysiological foundations of emotions, attachment, communication, and self-regulation. W. W. Norton & Company.
Schultz, W. (1997). Predictive reward signal of dopamine neurons. Journal of Neurophysiology, 80(1), 1–27.
Schultz, W. (2016). Dopamine reward prediction error coding. Dialogues in Clinical Neuroscience, 18(1), 23–32.
Schulz, A., & Vögele, C. (2015). Interoception and stress. Frontiers in Psychology, 6, 993.
Witkiewitz, K., Wilson, A. D., Pearson, M. R., Montes, K. S., Kirouac, M., Roos, C. R., Hallgren, K. A., & Maisto, S. A. (2019). Profiles of recovery from alcohol use disorder at three years following treatment: Can the definition of recovery be extended to include high functioning heavy drinkers? Addiction, 114(1), 69–80.